Small, hydrophobic molecules can pass hydrophobic membrane easily without channels. Small, uncharged polar molecules can also pass but less easily; large, uncharged polar molecules and ions almost cannot pass membrane freely.
1. Independent of plasma membrane proteins
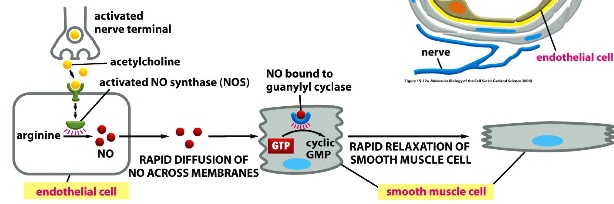
small molecule: Nitric Oxide (NO)
- made by deamination of arginine by NO synthase
- example of a short-range molecule because it has a short half life (5-10 seconds)
- acts on smooth muscle and other target cells
- smooth muscle cells wrap around endothelial cells that surround blood vessels
- nitroglycerine is converted to NO to reduce workload on the heart —> alleviate heart pain
***activated nerve termini send out vesicles with acetylcholine, which then binds to a receptor on the endothelial cell surface —> activates NO synthase which then makes NO from arginine —>NO rapidly diffuse out of endothelial cells and into smooth muscle cells —> NO binds to guanylyl cyclase, changes its conformation —> signal is sent to convert GTP into cGMP —> cGMP leads to the relaxation of smooth muscle cells
Small hydrophobic signalling molecules
steriod hormones — made from cholesterol
thydroid hormones — made from tyrosine
retinoids — made form Vitamin A
Vitamin D — affects metabolism
^ be familiar with some examples, not details
Important to know is that these molecules, since they are hydrophobic, cannot be transported in the blood stream without carrier proteins. Once they arrive at their designated cells, they need to dissociate from carriers and enter the cell where they can bind a member of the nuclear receptor superfamily (but not necessarily INSIDE the nucleus, but when they bind to the ligand, they move into the nucleus to change gene expression).
Nuclear receptor superfamily
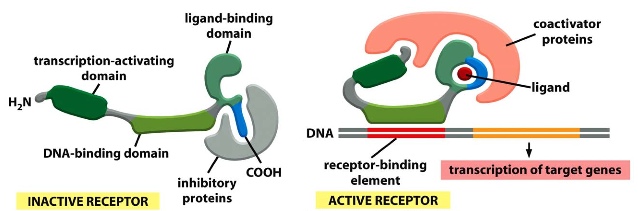
- has a binding site for a small hydrophobic molecule and DNA
- 48 of nuclear receptors members are identified in the human genome
- we know what the small ligand is for more than half of the nuclear receptors, but for those ligands that are unknown, they are called orphan nuclear receptors
- when it is inactive, the ligand-binding domain is covered up by an inhibitory protein, when ligand binds to the active site, the inhibitory protein is released, the receptor conformation changes and a coactivator protein covered transcription-activating domain and ligand-binding domain —> complex binds to receptor-binding element—> result is transcription
The transcriptional response to hormones can have many steps:
Primary response: receptor + ligand acts on DNA and leads to transcription of certain genes —> synthesis of proteins after translation
Secondary response: primary response proteins act on DNA to inhibit or promote translation of other genes and thus other proteins.
Secondary can activate tertiary … etc.
2. Through plasma membrane channels
ion channel
- there are many functions, but mainly electrical excitability of muscle cells and signalling in the nervous system.
- they have narrow pores so that only a selective group of ions can pass through
- close and open rapidly and allow many many ions to pass through at the same time
- passive transportation — no ATP required, depends on the electrochemical gradients across the plasma membrane: moving from HIGH to LOW concentration
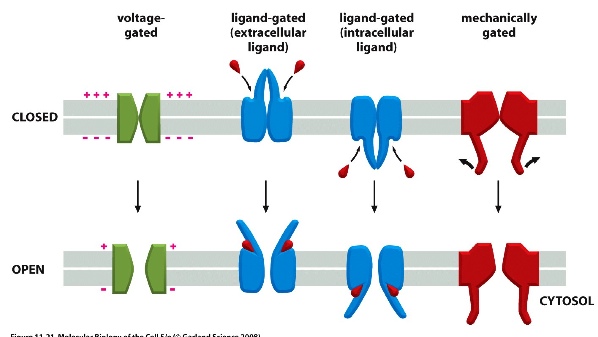
- main types: voltage-gated (usually more + charge on the outside of the cell); ligand-gated; and mechanically gated.
- Na+ tend to be found on the outside; K+ on the inside —> in a neutral cell, there should be a balance of charges
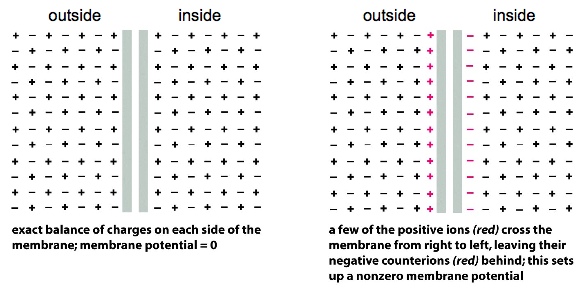
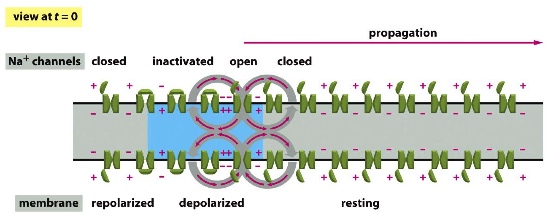
There’s an active pumping of 3 Na+ to the outside vs. 2 K+ to the inside, so that the outside of the cells is usually slightly more positive than the inside. This makes the membrane polarized. When the Na+ channels are open, Na+ are allowed to move inside, depolarizing the membrane. —> induces a travelling wave of depolarization (action potential)
- Closed channels: more + on the outside, polarized membrane, can be opened by depolarization of previous channels
- Open channels: Na+ going into the cell, making the inside more positive —> depolarization
- Inactivated channels: after the channels opening for a while, they will become inactivated so cannot be reactivated for a while until they are closed again and returned to the original state
Since the inactivated channels cannot be reopened again, the signal only moves towards the terminus and cannot move back, making a directional signal moving forward.
At the end of the presynaptic nerve terminal, neurotransmitter-container vesicles are released into the synaptic cleft —> neurotransmitters diffuse through and bind to the ligand-gated channels on the postsynaptic neurons.
3. Downstream of plasma membrane G-protein coupled receptors
G-protein coupled receptors(GPCR) signal via small molecules in more complicated ways.
> 7 pass transmembrane proteins
> activated by proteins, small molecules and light
> more than 700 in humans
> all of theses receptors signal into the cytosol via a membrane associated trimeric(alpha, beta, gamma) GTP-binding protein (G protein)
Alpha subunits can bind to GDP or GTP.
The activated receptor functions as a GEF which exchanges GDP for GTP on the a-subunnit —> conformational change —> releases b-subunit and c-subunit —> triggers downstream effectors. The activity is turned off by regulator of G protein signalling (RGS) which acts like a GAP(GTP—> GDP).
Sometimes only one subunit activates downstream effectors, sometimes all of them.
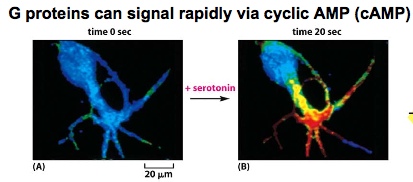
G Proteins signall rapidly via cyclic AMP. With serotonin added, the warm dye signals the presence of cAMP. cAMP is made from ATP by adenylyl cyclase and destroyed by cAMP phosphodiesterase.
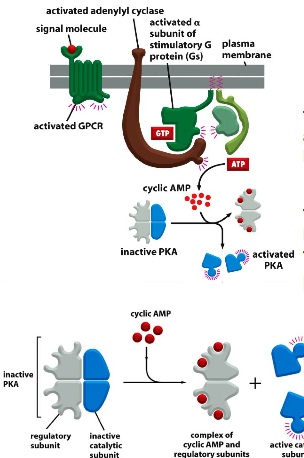 <important pathway>
<important pathway>signal molecule binds to G-protein coupled receptors —> activated GPCR acts like GEF and activates G-protein —> a-subunit releases b and c-subunites —> certain subunits activate adenylyl cyclase —> makes cAMP from ATP —> binds to the regulatory subunits of PKA and release the catalytic subunits —> phosphorylates downstream effectors.
(there’s constant background activity of phosphodiesterase to prevent accidental signalling. This basically establishes a threshold for the signal, so the right signal only comes through when there is an intentionally increased activity of adenylyl cyclase)
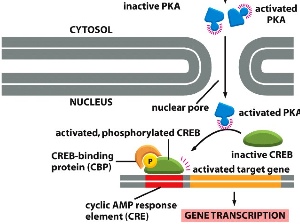
activated PKA phosphorylates inactive CREB —> CREB binds with CBP and PO4 —> binds to CRE on DNA and starts gene transcription.
e.g. cAMP signalling in sense of smell
> each receptor recognizes an idv. set of odourants —> cAMP —> opens on cAMP-gated cation channels—> action potentials (one odourant to one receptor: combination of different signals allow us to distinguish numerous different smells)
e.g. GPCR signalling via calcium
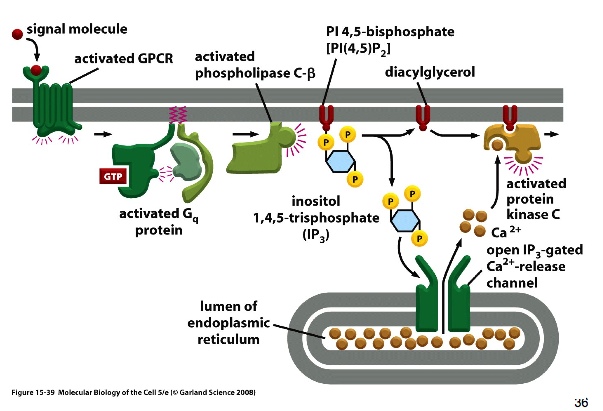
> signal activates GPCR —> G protein activated —> activates phospholipase C —> activates PIP2 to cleave it into two new signalling molecules: IP3 and DAG.
IP3 is water soluble and can diffuse through the cytosol; DAG is hydrophobic that diffuses along the plasma membrane.
IP3 binds to IP3 gated channels on the surface of ER (ER stores Ca2+) —> Ca2+ released into the cytosol; meanwhile, DAG diffuses through plasma membrane and activates protein kinase c (PKC) —> Ca2+ binds to PKC and activates it —> phosphorylates downstream targets.
- Ca2+ not only binds to PKC but also other targets, as well as DAG
- PKC is a serine/threonine kinase